与真核生物的细胞核不同,原核生物的遗传中枢是非膜结构的“拟核”,遗传物质及其相关蛋白质共存其中。在近十年对各种原核生物的高级染色质结构研究中,最受关注的方向包括:拟核相关蛋白(NAP)对建立和维持全局或局部染色质结构的影响、细胞周期进程中染色质结构的动态变化和染色质结构维持复合体(SMC)对这一过程的作用,以及SMC协同ParABS系统在细胞增殖过程中保证染色质结构维持和分离过程的机制。链霉菌(Streptomyces)基因组上存在多种天然产物生物合成基因簇(BGC),其负责合成多种用于临床和农业防治的生物活性小分子化合物。然而,随着多种临床耐药菌的出现,发掘具有医用价值的新化合物或者提高现有高应用价值天然产物的产量显得尤为重要。目前高产天然产物的方法主要包括外在培养条件的优化和一维水平下内在代谢工程改造等。然而,因为原核生物中基因表达和染色质结构之间的潜在联系仍远未被完全揭示,如何在三维水平上,利用染色质高级结构特征来提高天然产物产量仍鲜有研究涉足。
该研究应用高分辨率的Hi-C方法对天蓝色链霉菌(S. coelicolor)A3(2)这一模式菌株在两种代谢特征的生长时期的染色质三维结构进行解析,考察了多类型的染色质三维结构特征,包括拟核互作结构域(CID)的鉴定和动态变化、CID边界基因特征分析和功能鉴定、基因水平转移区的显著互作变化、遗传分区与结构分区之间的不对应性等。同时发现局部染色质的空间重排与BGC在不同生长时期的表达变化有关。接着,该研究发现内源基因的转录水平与局部染色质互作频率(文中量化为FIRE值)高度相关。然后,将一个编码β-葡萄糖苷酶的报告基因gus(a)和一个合成天然产物RK-682 的BGC插入不同的基因组环境时,gus(a)的表达量和RK-682的产量与FIRE值都成显著正相关。这说明无论是内源还是外源的基因、单个基因或包含一组基因的基因簇,染色质局部互作频率都将显著影响它们的表达水平。最终,该研究应用这一规律实现了相对于传统方法的天然产物高产。
1、S. coelicolor的高级染色质结构
BGC的表达水平伴随着不同生长阶段发生转变(在指数生长中期(M期)之前几乎沉默,在指数生长后期(L期)之后高表达)。该研究通过Hi-C追踪S. coelicolor在两个生长时期的基因组三维结构变化是否适应这一生物学过程。
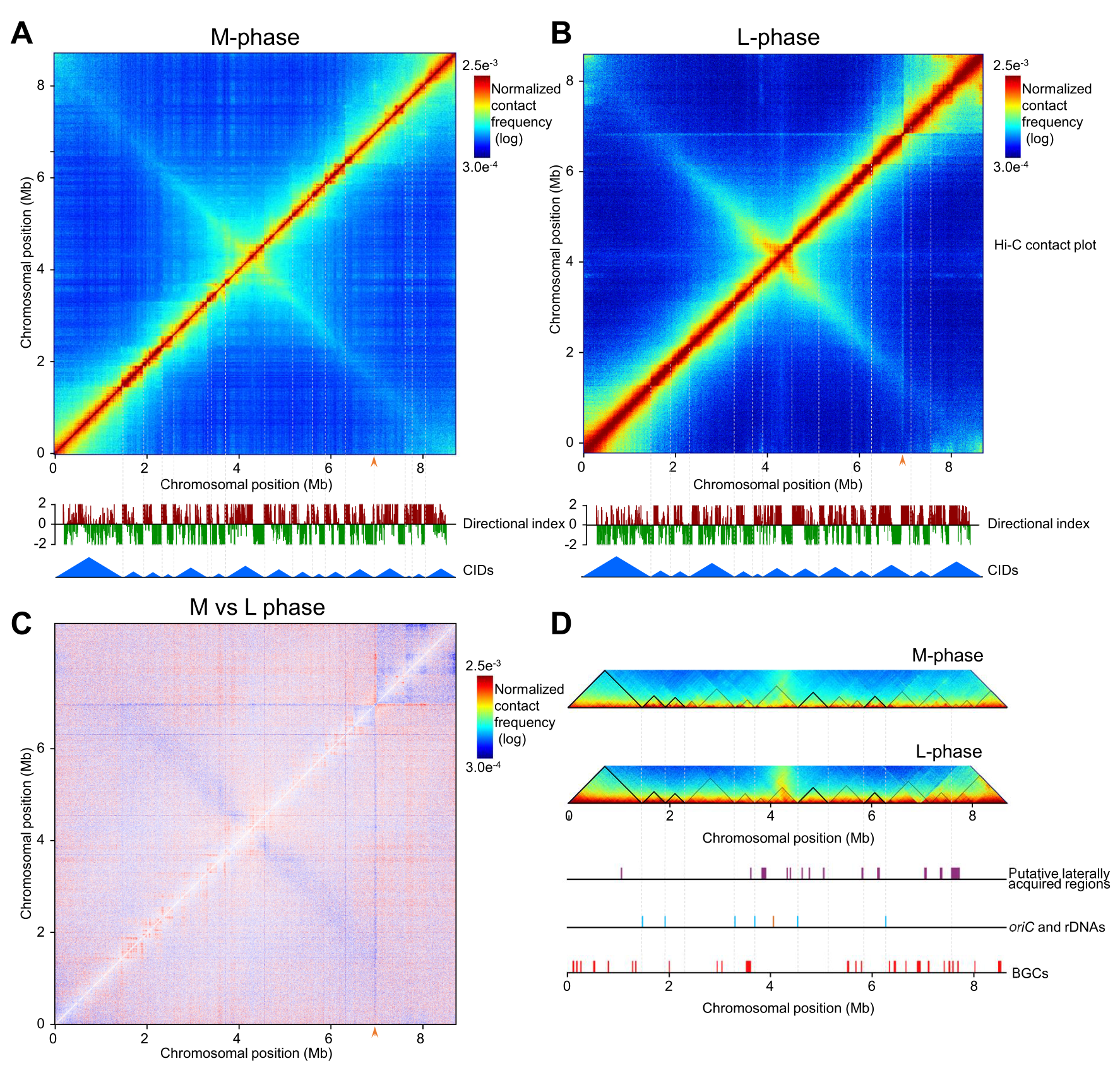
首先,两个时期的全局互作图都展现出一条主对角线,代表临近基因区的强互作,还有一条与主对角线垂直的次对角线,从复制起点(oriC)附近延伸到染色质的所有其他区域。次对角线的宽度和颜色饱和度随着时间的推移而增加,表明随着生长时期转换染色质发生了一定程度的压缩。同时,线性染色质的末端区存在弱的互作信号,这印证了前人关于末端蛋白的共价结合使基因组实现遗传连锁的细胞学实验结果。
按照基因保守性不同,天蓝色链霉菌基因组可以分为一个居中的核心区和两端的染色质臂区。利用Hi-C数据划分染色质compartment和3D结构可视化后,发现遗传分区和三维结构分区之间存在部分不协调(整个左臂和核心区域的一小部分一起构成了左侧compartment B)。
之后,对于L期互作矩阵的异常线条进行了分析,4.3 Mb处的线条显示了ori区域与染色质其余部分之间的相互作用。这可能是ParB-oriC分离复合体沿拟核的分离和移位引起。6.9 Mb处的线意味着该区域与染色质的其余部分之间的全局和明显的相互作用。该区域包括三个前体供应基因(SCO6269~SCO6271),负责合成丙二酰-CoA这一聚酮化合物的延伸单元,以及两个与BGC转录调控相关的γ-丁酰内酯信号(GBL)通路相关的调控基因(SCO6265~SCO6266),这暗示了天然产物生物合成与染色质互作之间的关联。
图1 M期(A)和L期(B)的互作矩阵,差减矩阵(C),CID差异(D)展示
在S. coelicolor中也存在染色质相互作用区域(CID,类似于真核生物中的TAD)。其中M期有17个CID(85~1470 kb),L期有13个CID(220~1470 kb)。虽然两个时期只有5个完整的CID完全相同,但仍存在10个相同的边界,说明CID在生长时期转换过程中的适应性结构调整(一个CID拆分为两个或多个较小的CIDs,相邻CID融合为一个较大的CID)。在M期和L期,绝大部分CID边界的GC含量低于染色质平均值,并且边界区存在许多已知转录因子和NAP的结合基序(motif),暗示DNA碱基偏好和相关因子对CID边界形成和维持的作用。
结合转录组数据分析发现,M期和L期CID有50%和56.6%的边界基因的表达水平较高,并且CID边界基因的转录强度高于内部基因。在CID边界,相当一部分高表达基因与细胞生长和发育所需的物质和能量代谢相关,并且高表达基因的转录方向和基因组复制方向之间有明显的一致性,而边界定位的低表达基因转录方向与基因组复制方向相同的趋势明显下降,因此基因的高表达可能是其转录方向与基因组复制方向趋同进化的结果。同时CID边界之间的显著相互作用从M期到L期明显减少。类似的现象也发生在14个鉴定的水平基因转移区中。在不同的生长阶段,这些位点之间的共定位和分散暗示了一种适应代谢状况过渡的调控机制。
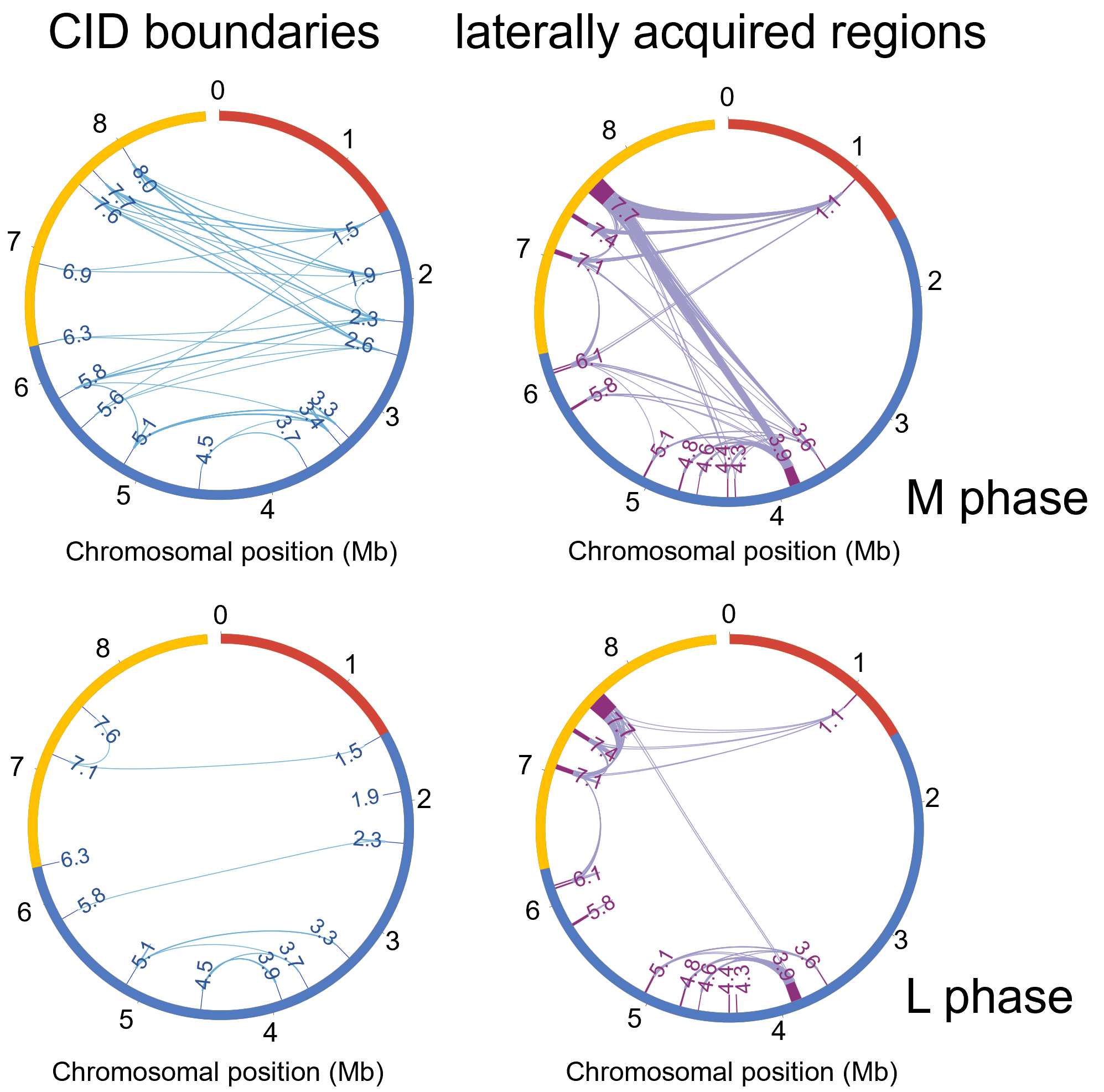
图2 M期(上)和L期(下)中CID边界之间和14个水平基因转移区之间的互作
2、天然产物生物合成相关局部染色质重构
为了研究代谢分化过程中天然产物生物合成和局部染色质三维结构重排之间的相关性,对天蓝色链霉菌基因组中所有28个BGCs在两个生长时期的局部染色质三维结构和转录水平变化进行分析,其中在2个BGCs里发现了两者之间的同步变化。
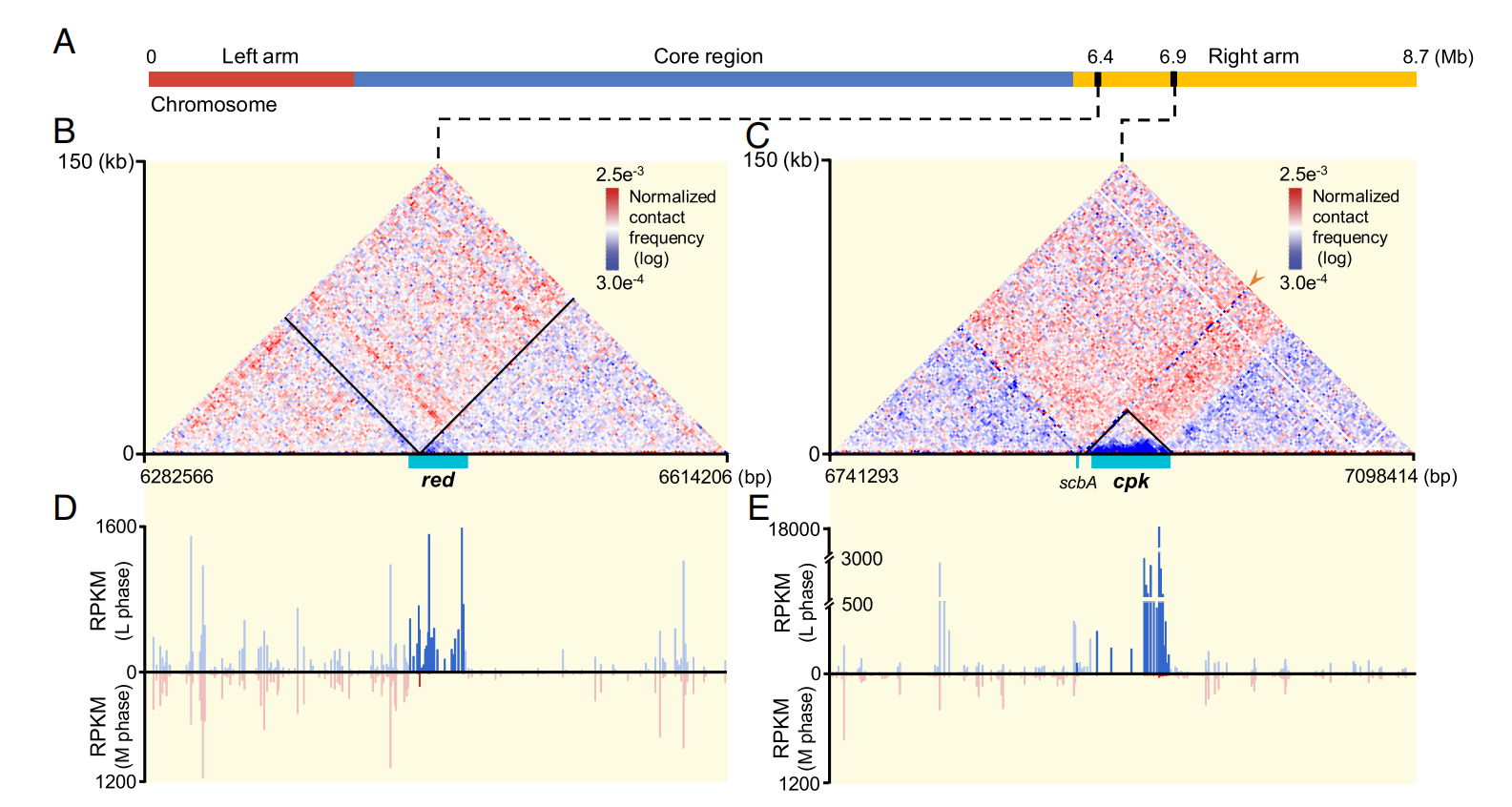
十一烷基灵菌红素是一种被广泛研究的红色色素。负责其生物合成的red BGC的转录水平从M期到L期增加了143倍,并且red重新定位于两个嵌套结构域(nested domain)之间。这主要是由于局部高转录对染色质结构域扩散的抑制作用所致,展现了转录对染色质结构的塑型作用。
负责天蓝霉素P1生物合成的cpk BGC的转录强度从M期到L期急剧增加541倍。惊奇的是,在L期观察到一个新形成的横跨整个簇的独立拓扑域。这与拟南芥中thalianol BGC的情况类似,反映了原核生物和真核生物间基因表达的三维调控机制的相似性。同时,这个拓扑域的上边界即是位于6.9 Mb基因座上,L期与全基因组存在互作的特殊DNA区域。这从一个更高的维度揭示出了一种全新的次生代谢调控机制,它表明链霉菌中天然产物的生物合成与染色质高级结构存在紧密联系,为其将来可能的应用提供重要的理论基础。对这一现象可能的机制模型和生物学意义分析,在文中进行了详细描述。
图3 red和cpk BGC的转录强度和局部染色质结构的协同变化
3、染色质局部互作频率显著影响内源基因和外源基因的表达水平
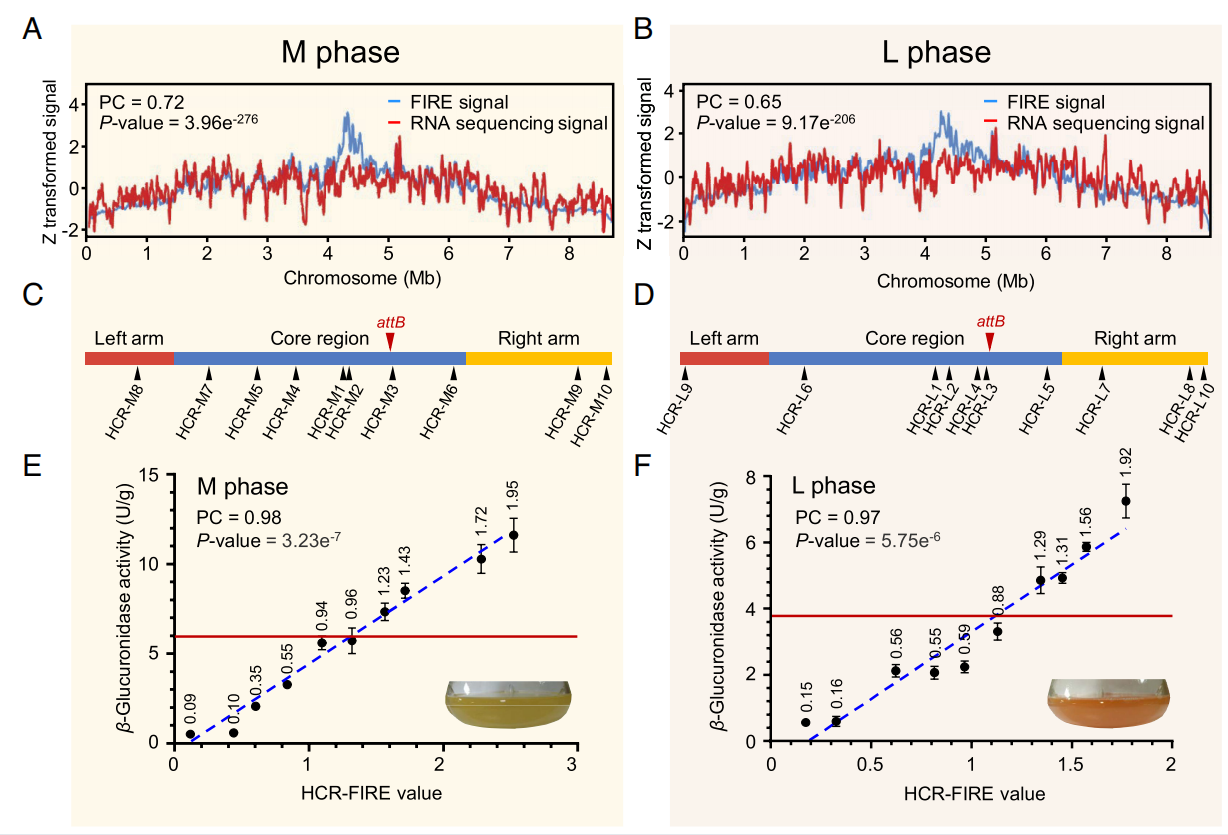
首先,5kb 分辨率的FIRE值(每个bin内局部顺式互作)和转录表达之间的显著相关明确了内在基因表达和局部染色质互作频率的紧密联系。为了探究染色质局部互作频率是否能直接影响该区域基因的表达,该研究首次将FIRE值与内在基因转录水平相关性大于0.8的染色质区定义为高相关性区域(High correlative region, HCR),然后按等分原则分别选定了FIRE值梯度变化的10个HCR区域(M期, HCR-M1~10;L期,HCR-L1~10)。接着将gus(a)报告基因分别插入这20个HCR区域,并分别测定报告基因表达强度。结果显示,在两个时期,报告基因表达强度与FIRE值都高度相关,并且表达量差异最高可达22倍。与整合到attB位点的传统方法相比,该报告基因的表达量最高可实现约2倍的提升。
图4 内在基因和gus(a)报告基因表达量和局部染色质互作频率的相关性
4、利用高FIRE值的HCR区实现天然产物高产
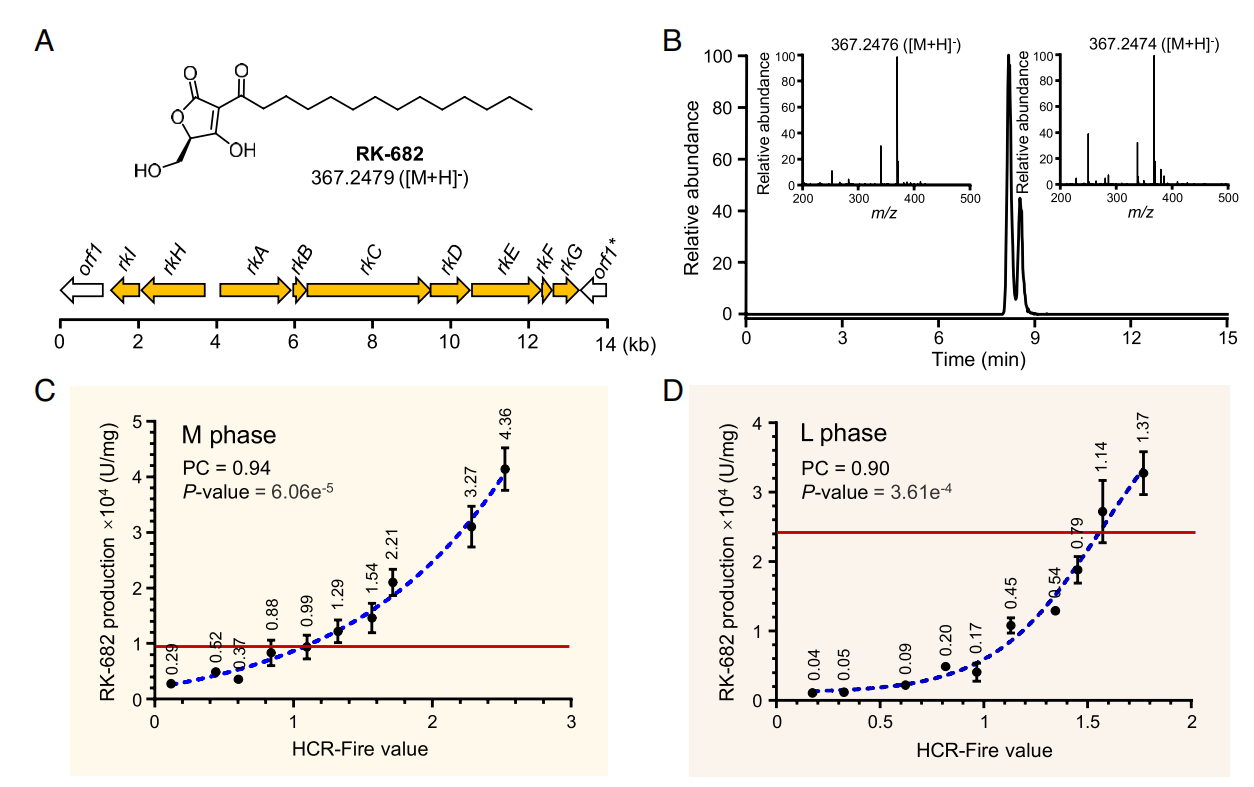
基于报告基因的积极结果,该研究进一步探索利用这一发现实现天然产物高产的可行性。将链霉菌Streptomyces sp. 88-682中负责天然产物RK-682生物合成的一个11.8 kb的BGC分别插入这20个HCR区域后,进行同步发酵和RK-682产量检测。结果显示,在两个生长时期,RK-682产量与FIRE值同样高度相关,并且表达量差异最高可达30倍之多。与整合到attB位点的传统方法相比,在高FIRE值HCR区域上的产量提升最高可达到4.3倍。这些结果不仅证明了染色质局部互作频率可以显著影响置入其中的内源或外源基因或基因簇的表达,并且利用这一发现实现了天然产物的高产。
图5 RK-682产量和局部染色质互作频率的相关性
这项工作展示了链霉菌在不同生长周期的适应性染色质结构动态变化和代谢转换,描述了染色质局部结构和天然产物生物合成基因簇转录的相关性,发现了局部染色质互作频率与基因表达之间的重要联系,并首先将这一发现付诸实践,在极具应用价值的链霉菌中实现了天然产物的高产。该研究将为原核生物的染色质结构和转录调控提供了新的见解,为高产天然产物提供了新的思路。
武汉大学邓梁、北京生物技术研究所赵志虎研究员为本文共同第一作者,武汉大学孙宇辉教授为通讯作者。5848vip威尼斯电子游戏刘琳、周帆署名参与了本研究。该研究得到了国家重点研发计划的资助。
 027-87224696
|
027-87224696
| marketing@frasergen.com
|
marketing@frasergen.com
|



 微信公众号
微信公众号

